Underwing in shorebirds: a global pattern of geographical variation
Arctic shorebirds are textbook examples of fragmented, discrete geographical variation — subspecies structured into populations separated by thousands of kilometres, diverging in measurements and plumage across glacially isolated breeding grounds. The pattern is well documented in circumpolar species such as the Whimbrel (Numenius phaeopus, with — among others — the nominate subspecies across Europe and most of Asia, variegatus in the Far East, and the recently elevated hudsonius in North America) and the Bar-tailed Godwit (Limosa lapponica, with the similar lapponica, yamalensis and taymyrensis across Eurasia, menzbieri in the Far East, and baueri in Alaska). In both cases, this geographical differentiation traces back to the isolation of breeding populations during Pleistocene glaciations — populations locked into separate refugia, diverging quietly over tens of thousands of years.
What makes these two species worth a second look, however, is not their clinal gradient per se, but the direction it has taken. In both species, Western Eurasian birds tend to have clean white backs and pale, unmarked underwings; Far Eastern birds are heavily barred; and North American populations are so densely barred that their underwings and rump appear uniformly dark in the field. A coincidence, perhaps — except that the same gradient appears, independently, in the European and Pacific/American Golden Plover group, in the Caspian/Oriental Plover complex, in the Red Knot, and even in Common/Wilson's Snipes. And when you step back further and compare species found exclusively on one side of the Atlantic — Eurasian Curlew and Black-tailed Godwit on the one hand; Long-billed Curlew, Hudsonian Godwit, Marbled Godwit, and Willet on the other — the contrast becomes almost schematic: white underwings in Europe, dark underwings in the Americas. This post argues that this is not a coincidence, and proposes a first hypothesis for why: that large shorebirds sharing a migratory flyway have been under selection to develop a similar underwing pattern, facilitating visual recognition within mixed-species flocks.
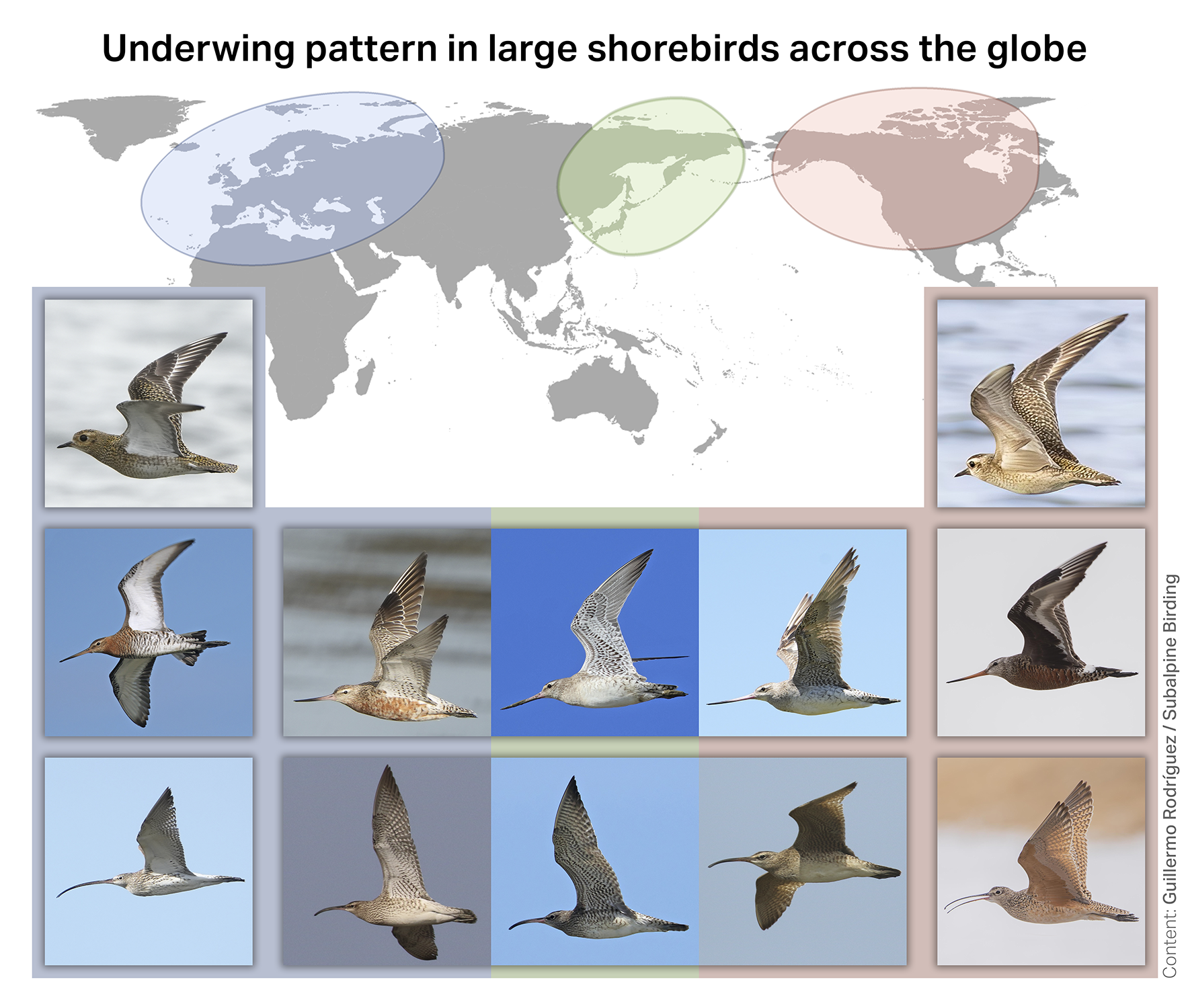
Plate 1. Underwing pattern in large shorebirds across the Northern Hemisphere, illustrating a global interspecific pattern of geographical variation. Left: species breeding exclusively in Europe or across Eurasia show white underwings — from top to bottom: European Golden Plover, Black-tailed Godwit, and Eurasian Curlew. Right: species breeding exclusively in the Americas show uniformly dark underwings — American Golden Plover, Hudsonian Godwit, and Long-billed Curlew. Centre: circumpolar species found across the entire Northern Hemisphere — Bar-tailed Godwit (top) and Whimbrel (bottom) — show barred underwings, but with a clinal gradient: unbarred or lightly barred in western Eurasia, becoming progressively denser until the underwing appears uniformly dark in their North American populations. Photo credits, by species: Hudsonian Godwit and Long-billed Curlew (Walker Kele), American Golden Plover (Gonzalo Pardo), Black-tailed Godwit, Asian Bar-tailed Godwit and Whimbrel (Dave Bakewell), rest of pictures (Guillermo Rodríguez).

Plate 2. Shorebird flocks appear predominantly white from below on the wintering grounds of Europe and Africa (Bar-tailed Godwits in S Spain, photo by José Manuel Reyes). In East Asia, most species look heavily barred — and the assemblage at any given location appears remarkably uniform in barring density, even though the overall intensity shifts as you move along the flyway (Bar-tailed Godwits and Whimbrels in Malaysia, by Dave Bakewell). In the Americas, by contrast, flocks of large shorebirds in flight present a vividly rich palette: the uniformly dark underwings of Willets combine with the warm orange-buff of Marbled Godwits and Numenius (flock in Virginia, by Mario Balitbit) — and, remarkably, Marbled Goodwit and Long-billed Curlew, the largest species among the Nearctic assemblage, show a strikingly similar underwing pattern (photo by Chris Wood).
Species overview
The pattern across groups
Summary tables displaying underwing and rump/back patterns across regions are presented below, scoring each species as pale, thickly barred, or dark across its geographical range. The west-to-east-to-Nearctic gradient — white or near-white in Europe and western Asia, heavily barred in the Far East, and uniformly dark in North America — is consistent and visually striking in the underwing, somewhat less so in the rump and back pattern. Critically, this gradient applies only to relatively large-bodied species (Red Knot-sized and above): godwits, Numenius curlews, Willet, and the larger plovers — precisely the species that characteristically associate in mixed-species flocks during migration and on wintering grounds. Small shorebirds — most Calidris sandpipers and Charadrius plovers — retain pale or white underwings regardless of their geographical origin, showing no equivalent biogeographic signal. Among the Tringa shanks, which are intermediate in body size, the pattern is present but less pronounced than in the larger species, suggesting a possible body-size threshold below which the selective pressures driving this gradient are either absent or insufficient to produce a consistent signal.
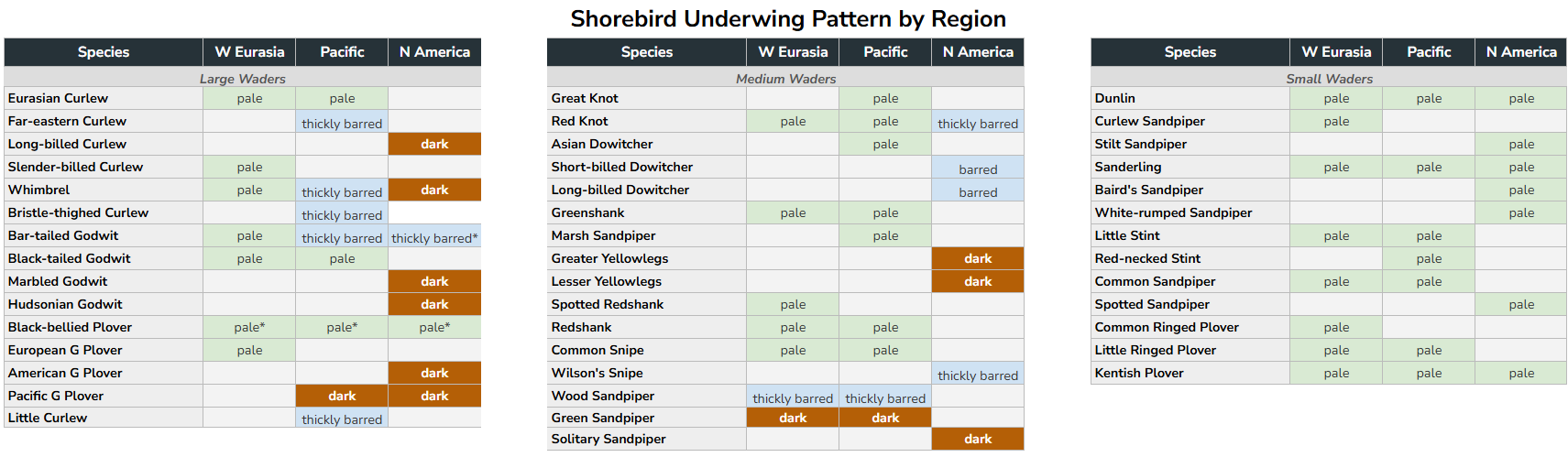
Table 1. Shorebird underwing pattern by breeding region, roughly ordered by size. Underwing patterns have been condensed into three categories: pale (overall pale, even if finely barred); thickly barred (barring extensive enough to produce a distinctly dark gestalt); and dark (uniformly non-white, typically brownish). Small shorebirds show no consistent geographic signal; large shorebirds show a clear west-to-east gradient from pale underwings in western Eurasia to dark underwings in North America. In medium-sized shorebirds the pattern is present but less well defined.
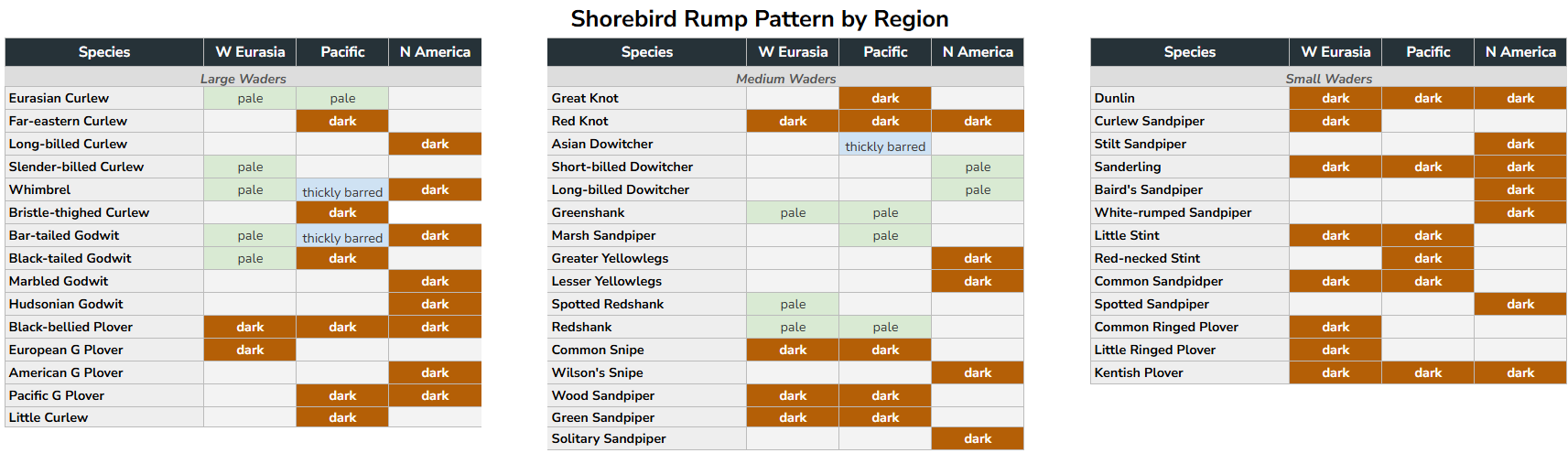
Table 2. Shorebird rump and back pattern by breeding region, roughly ordered by size. The same three categories apply as in Table 1. The geographic gradient is less consistent than in the underwing, and the pattern is largely driven by the pale, unmarked backs and rumps of western Eurasian representatives of key groups — particularly Numenius curlews, Bar-tailed Godwit, and the larger Tringa species — contrasting with the more heavily marked or uniformly dark rumps of their Pacific and Nearctic counterparts. In smaller shorebirds the rump pattern shows no meaningful geographic signal.
Numenius: the gradient is perhaps most unambiguous in this genus. All European and western Asian species — Eurasian Curlew, Whimbrel, and Slender-billed Curlew — present clean white underwings, while the West Pacific / Alleutian species — Bristle-thighed, Far Eastern, and Little Curlew — display dark but non-uniform, barred underwings. The sole truly extant Nearctic representative, Long-billed Curlew, shows uniformly dark underwings. Based on the few available photographs of museum skins, the Eskimo Curlew appears to have presented a uniformly warm orange-buff underwing similar to that of the Long-billed Curlew, albeit with more extensive but fine barring. As noted above, the Whimbrel — the only species spanning all three regions — exhibits a clear clinal gradient in underwing barring intensity from west to east. The Eurasian Curlew provides an instructive contrast: distributed across Europe and Asia but absent from the Americas, it shows no equivalent gradient, or arguably a reversed one, with western European birds frequently showing barred underwings while Asian individuals consistently present clean white underwings — a counterintuitive pattern that merits further attention.
Limosa: the Black-tailed Godwit, represented by several subspecies distributed across Europe and Asia, invariably presents unbarred white underwings regardless of population. Both Nearctic godwit species — Hudsonian and Marbled — display uniformly dark underwings. The Bar-tailed Godwit, the only species spanning all three regions, mirrors the Whimbrel in showing a pronounced clinal gradient in underwing barring intensity, reinforcing the impression that circumpolar species occupying all three migratory flyway regions independently recapitulate the broader interspecific pattern.
Pluvialis Golden Plovers: one of the primary identification criteria distinguishing the European Golden Plover from the Pacific and American Golden Plovers is, precisely, underwing coloration: pure white in the former, uniformly brownish in the latter species pair. Pacific and American Golden Plovers are more closely related to each other than either is to the European, having diverged more recently — yet molecular estimates place their separation at approximately 1.8 million years ago (Withrow & Winker, 2014), a timeframe more than sufficient for the acquisition of a distinctively paler underwing in one lineage, had selective pressure existed to drive it.
Tringa and allies: the Willet stands apart within this group, presenting a strikingly dark underwing consistent with the broader Nearctic pattern. The remaining large Tringa species — yellowlegs, redshanks, and greenshanks — do not exhibit a well-marked Nearctic/Palearctic difference in underwing coloration per se, though the contrast is conspicuous in rump pattern: yellowlegs show a dark rump, consistent with other large Nearctic waders, while Palearctic Tringa species characteristically display a pale rump and uppertail. Whether yellowlegs average more heavily barred underwings than Common Greenshank is plausible but would require a more quantitative analysis to confirm. Both redshank species (T. totanus and T. erythropus) present clean white underwings. An intriguing exception within this group is the Tringa sub-clade comprising Green, Wood, and Solitary Sandpipers — all three show distinctly dark underwings irrespective of their Nearctic or Palearctic distribution, suggesting that in this case underwing darkness may reflect shared ancestry rather than biogeographic signal. This is consistent with their unusual ecology among Tringa: all three are tree-nesting species that in winter/migration are strictly associated with freshwater habitats compared to their more coastal, long-distance congeners, raising the possibility that they were not exposed to the same selective pressures.
Anarhynchus Caspian and Oriental Plovers: this species pair is particularly instructive, since despite their relatively small body size — which elsewhere in this account correlates with absence of the pattern — both align well with the geographical gradient. The two species breed allopatrically across the Central Asian steppes — Caspian Plover in Kazakhstan, Oriental Plover in Mongolia — yet their migratory trajectories diverge sharply: the Caspian Plover heads southwest towards South Africa, and presents a clean white underwing accordingly; the Oriental Plover heads towards Australia, joining the Pacific flyway assemblage of barred and dark-underwing species — including Pacific Golden Plover and Little Curlew — and presents a uniformly dark brown underwing. Did the ancestral Anarhynchus population's adoption of divergent migratory routes — and the consequent differential association with Palearctic versus Pacific shorebird assemblages — drive not only the divergence in underwing coloration but ultimately the speciation event itself?

Plate 3. Underwing pattern in the genus Numenius: European (a) and Siberian Whimbrels (b) illustrating the clinal difference in barring intensity, mirrored by the underwings of the Eurasian and Far Eastern Curlews (e). Little Curlew (c) shows a similar underwing to that of the Nearctic Bristle-thighed Curlew (g) and Hudsonian Whimbrel (d), although in these two the background colour is warmer and richer — though still not approaching the deep orange-buff of the Long-billed Curlew (h). Photo credits: Guillermo Rodríguez (a, c); Dave Bakewell (b, e); and Alan Kneidel (d, g, h).

Plate 4. Caspian Plover (Ian Reid) and Oriental Plover (Guillermo Rodríguez) are sister species whose identification in non-breeding plumage can be challenging — until they show their underwing. Was the dark underwing of the Oriental Plover the mechanism that steered the migration of eastern populations of their common ancestor towards Australia, following Pacific Golden Plovers and Little Curlews migrating through Mongolia?

Plate 5. Pacific Golden Plover (Dave Bakewell) and American Golden Plover (Daniel López-Velasco). Based on the literature, the Pacific Golden Plover diverged from a Nearctic ancestor long ago — yet has retained a dark underwing. In many photographs, Pacific Golden Plover appears to show a subtle contrast between the axillaries and the remaining underwing coverts, suggesting a slight lightening of the latter; American Golden Plover, by contrast, tends to display a more uniform underwing. This difference is however difficult to assess reliably from photographs due to varying light conditions, and would require a more systematic study to confirm.
Circumpolar species as case-studies
Whimbrel
As described above, the Whimbrel perhaps represents the most paradigmatic example of the global pattern of geographical variation. Birds from the western part of Eurasia are finely barred, with both underwing and rump appearing essentially pale at distance. The subspecies migrating along the Pacific flyway are much more heavily barred — with thick bars as wide as or wider than the pale interspaces — and the rump is strongly patterned. In the Hudsonian Whimbrel, which breeds in boreal North America and overwinters in Central and South America, the back feathers are uniformly dark and unpatterned, and the underwing feathers are not only heavily barred but the background colour is already buffy/sandy rather than white — having already closely approached the appearance of other Nearctic species.
It is worth examining the pattern of Whimbrel in Europe and Central Asia in more detail, since the gradient appears locally reversed here. The palest underwing is shown by the Steppe Whimbrel, an isolated subspecies breeding in Central Asia. The nominate subspecies breeds further north and shows considerable individual variation in underwing tone, ranging from entirely white to consistently barred. The Icelandic population, however, tends to be more frequently barred, producing a slightly darker underwing on average — far from the condition of Pacific birds, but with a statistically detectable shift in the character distribution. Interestingly, a parallel local reversal is found in Atlantic Eurasian Curlews, which show more extensive barring than their continental counterparts. It seems plausible that this locally reversed pattern is coupled between the westernmost populations of Whimbrel and Curlew — a point that merits further investigation.
Bar-tailed Godwit
Like the Whimbrel, the Bar-tailed Godwit represents a striking example of clinal variation. The western Eurasian subspecies — lapponica in Scandinavia, taymyrensis and yamalensis in northwestern Siberia, wintering in Europe, W Africa and the Middle East respectively — show variable barring ranging from pure white to slightly barred underwings. Birds overwintering in Southeast Asia (ssp. menzbieri) present much thicker bars on the underwing and a heavily marked rump; the Nearctic subspecies baueri, which winters in New Zealand, displays even denser barring on a white ground colour in the underwing coverts, but most notably the rump feathers are uniformly dark rather than barred. baueri has partially colonised the Nearctic, but its migration remains transoceanic across the Pacific, unlike the Hudsonian Whimbrel. Will Nearctic Godwits continue to darken their underwing? And if so, could that shift eventually redirect their migration towards a South American wintering strategy?
Red Knot
The Red Knot is phylogenetically a Calidris, but its large size means it regularly associates in flocks with larger shorebird species. It is well represented across all three regions, with a number of recognised subspecies — though clear field identification criteria to separate the different forms remain to be established:
canutus breeds in West Siberia and winters in West and South Africa.
islandica breeds in Greenland and winters in the British Isles and Western Europe.
piersmai and rogersi breed in two isolated populations in East Siberia, and winter mainly in Australia and New Zealand.
roselaari breeds in Alaska and winters in California.
rufa breeds in the Canadian Arctic and winters in Argentina.
Looking closely, there appear to be statistical differences in underwing barring among several of these subspecies (**): canutus presents the cleanest underwing, often looking entirely white; piersmai, rogersi, and roselaari present largely pale underwings, but with quite variable barring; rufa shows a considerably darker underwing due to dense barring; and islandica, although paler than rufa, often shows thick barring. This might, in principle, appear to break the rule of the global underwing pattern — particularly because islandica winters in western Europe, which would suggest a recent colonisation of Greenland from the continent; and similarly, one would expect roselaari to show a typically Nearctic dark underwing.
However, Conklin et al. (2022) used genome-wide SNP data to demonstrate that islandica and rufa — despite wintering on different continents — are virtually indistinguishable in neutral genetic variation, suggesting that islandica colonised its current Canadian and Greenlandic breeding grounds from a Nearctic refugium and only subsequently evolved its trans-Atlantic migration route to European wintering grounds, likely within the last few thousand years. Similarly, roselaari clusters with the East Asian taxa, suggesting it only recently colonised Alaska from a Siberian origin, as also supported by palaeoflyway reconstruction (Buehler et al. 2006).
Did the underwing pattern already carry a signal of each taxon's origin? It is interesting to note that, although identification criteria among Red Knot subspecies are still being developed, rufa and canutus are morphologically much more similar to each other — long-billed, robustly built, and grey above in breeding plumage (more on this in SubalpineBirding soon!) — than either is to islandica. It is possible that the demands of a long-distance, trans-tropical migration drove phenotypic convergence between rufa and canutus, while the underwing remained unchanged, retaining a signal of each taxon's true geographical origin.
(**) It is worth noting that Red Knots show considerably broader individual variation in underwing barring than the larger species discussed above — arguably a consequence of their intermediate size, which leads them to regularly associate with smaller Calidris in mixed flocks. If the selective pressure driving underwing convergence operates primarily through size-assortative flocking, Red Knots may simply sit below the threshold where that pressure is strong enough to produce a consistent signal.
Grey (Black-bellied) Plover
The Grey Plover is the only large shorebird that does not conform to the underwing darkness gradient: all populations across the globe show a clean white underwing with a sharply contrasting black axillary patch. Notably, the Grey Plover is also the only species with a truly distinctive underwing pattern that fits neither the plain white nor the plain dark categories shown by most other species in this account, and this singularity may itself be responsible for its non-conformity with the global pattern. Interestingly, where both species co-occur, Grey Plovers regularly associate in flocks with Dunlins — a small Calidris that shows white underwings regardless of geographic origin, and which therefore exerts no directional pressure towards darker underwing signalling.

Plate 6. Red Knot subspecies underwing variation: rufa (left, Kathy Horn), canutus (right, Guillermo Rodríguez), and islandica (bottom, James Kennerley). Note the striking difference in barring density between the three subspecies — and the unexpected similarity between islandica and the Nearctic rufa. There is considerable individual variation in barring within each subspecies, with islandica and canutus showing partial overlap between them.

Plate 7. Grey (Black-bellied) Plover. (Left, Guillermo Rodríguez) This species regularly associates with Dunlins in winter wherever both species coincide. Has the distinctive underwing pattern of the Grey Plover played a role in this unusual interspecific association? (Right, Jesse Pline) Look at this individual from Florida, US — are Nearctic Grey Plovers already exploring the advantages of darkening their underwing, as in other North American species?
Reading between bars
Just a coincidence?
Assessing whether patterns in biology reflect genuine evolutionary pressures or arose by chance is always challenging, as it generally requires strong — and often arbitrary — assumptions about the prior distributions driving such mechanisms. But let us attempt a simplified, naïve exercise to get a sense of the odds of the described geographical variation in underwing pattern arising purely by coincidence.
Assuming that the probability of a given large shorebird species having either an all-white or all-dark underwing is 0.5 — a conservative estimate, given that roughly half of all wader species show predominantly dark underwings — and that species within each region evolved their underwing independently of one another, then the probability of all 7 Nearctic species in the main groups described above showing dark underwings (American Golden Plover, Hudsonian and Marbled Godwits, Willet, Eskimo and Long-billed Curlews, and Hudsonian Whimbrel) is p_NA = 0.78%; the equivalent probability for the 6 European taxa (Black-tailed Godwit, Bar-tailed Godwit, Slender-billed and Eurasian Curlews, European Whimbrel, and European Golden Plover) all showing pale underwings is p_EUR = 1.56%; and the likelihood of both patterns occurring simultaneously by chance is 0.012%. It should be noted that this calculation assumes phylogenetic independence between species — an idealisation, since related species share evolutionary history. However, the fact that the pattern is consistent across multiple unrelated families (Scolopacidae and Charadriidae alike) provides at least partial support for the independence assumption. We can reasonably conclude that the geographical pattern is unlikely to be coincidental, and that some underlying mechanism must be at work.
The "grouping-by-underwing" hypothesis
The underwing pattern is undoubtedly important in shorebird communication, as many species actively display their underwings during social and sexual displays — making it straightforward to relate underwing variation to speciation mechanisms such as assortative mating. The fact that all species within a given region converge on a broadly similar underwing pattern is more intriguing, however, as it could reflect either a shared environmental pressure acting similarly across species, or alternatively a coupled speciation process. A purely environment-driven explanation seems unlikely: the pattern consistently involves species breeding across a wide range of latitudes, longitudes, and ecological contexts, with very different feeding techniques and habitat preferences.
What these species do share is their migratory routes and roosting sites. The most parsimonious hypothesis is therefore that large shorebirds sharing a migratory flyway somehow benefit from having a similar underwing pattern, even if the precise mechanism remains to be demonstrated. One possibility is a selective pressure operating across large species: for instance, helping inexperienced juvenile birds to associate with heterospecifics on the same flyway and navigate towards the correct breeding or wintering grounds, even when separated from conspecifics. Alternatively, it could operate as a selective mechanism at the flock level: imagine a mixed-species shorebird flock on an American mudflat threatened by a Peregrine Falcon — all birds flush simultaneously, and a large dark-underwing species inadvertently joins a flock of small pale Calidris, becoming a conspicuous and disproportionately targeted outlier. A shared underwing pattern across large species would help birds sort by size and appearance during such events, homogenising flock composition and complicating prey selection by the predator. These are illustrative examples rather than fully developed hypotheses, and many other mechanisms are plausible.
The idea of coupled evolution between species sharing a migratory pathway has previously been proposed for American wood warblers by Gayk et al. (2021), who documented acoustic convergence between species with similar migratory routes and proposed selection for interspecific flock cohesion during nocturnal migration as the driving mechanism — essentially a communication channel maintained across species lines. What we propose here is conceptually similar, with visual rather than acoustic signals as the currency of that cross-species channel.

Plate 8. A Nearctic Bar-tailed Godwit, ssp. baueri, blending remarkably well into a flock of uniformly dark Marbled Godwits (Skip Cantrell). Note that while baueri has acquired uniformly dark uppertail coverts — like other Nearctic species — the rump feathers remain heavily barred rather than fully dark, suggesting this population sits at an intermediate stage of the darkening process. Compare with the Far Eastern menzbieri, in which the back feathers are barred rather than uniform.

Plate 9. A vagrant Hudsonian Godwit in the UK, associating with a flock of Black-tailed Godwits (Simon Colenutt). Once considered a single species, the two are immediately told apart in flight — their underwings could hardly be more different.
How did it all start?
The hypothesis outlined above postulates that large shorebirds adjust their underwing covert pattern to converge on that of co-migrants along their shared flyway. What it does not explain is who, or what, initiated the distinction in the first place. Was there a founder effect in a single species, with others subsequently following? Is the Nearctic environment intrinsically more conducive to the evolution of dark underwings, perhaps through some aspect of light environment, predator community, or ancestral plumage state? These questions remain open for future exploration.
There are some interesting cases, however, worth examining, as they may reveal traces of the evolutionary trajectories involved. The genus Numenius is, as mentioned earlier, quite paradigmatic — its high diversity in the Palearctic (five strictly Palearctic species, one Pacific/Aleutian, one circumpolar, and two strictly Nearctic) strongly suggests that the genus originated there. The Long-billed Curlew shows an underwing colour extremely similar to that of the Marbled Godwit, and a small proportion of individuals retain a remnant barring in the axillaries. The Hudsonian Whimbrel's underwing is similar in tone, but the barring remains very thick — comparable to that of the Eurasian Whimbrel subspecies variegatus. It is tempting to postulate that the ancestral Long-billed Curlew colonised the Nearctic and progressively coupled its underwing signal with that of the Marbled Godwit; and that the Hudsonian Whimbrel did the same during a later, independent colonisation of the region.
In the opposite direction, the Pacific Golden Plover likely had a Nearctic origin, and as mentioned above genetic studies dated its divergence from the American Golden Plover to approximately 1.8 million years ago. Despite this deep evolutionary separation, the Pacific Golden Plover has retained a dark underwing — even if, as discussed in Plate 5, it may have lightened slightly. Perhaps the dark underwing remains well-suited to the Pacific shorebird assemblage, in which most species show thickly barred underwings that are dark enough to remain compatible with a uniformly dark signal. At the same time, it seems intuitive that the transition from uniform dark back to barred does not occur readily — the evolutionary ratchet, in this case, may only turn in one direction.
Feather barring as a tuneable phenotypic dial
One of the most remarkable aspects of this pattern is how smoothly graded it is — not a discrete switch between dark and pale, but a continuous cline of barring density from clean white through progressively thicker bars to uniform darkness. This gradience has an interesting genetic parallel. Research in chickens has shown that feather barring is controlled by a surprisingly small number of genes — principally MC1R, which promotes dark pigment production, and its antagonist ASIP, which suppresses it. Small mutations in either gene, or in the regulatory sequences that control when and where they are expressed during feather growth, are sufficient to shift a feather from unbarred to heavily barred or uniformly dark. The within-feather periodic pattern itself — alternating dark and pale bands — is thought to arise from a Turing-type reaction-diffusion process: the two molecules activate and inhibit each other across the growing feather in a self-organising wave, producing bars whose width and spacing are set by the relative diffusion rates of activator and inhibitor.
Crucially, this kind of system may be easy to tune quantitatively: a modest shift in the balance between MC1R and ASIP expression can smoothly increase or decrease bar density without requiring a wholesale genetic overhaul. It seems likely, therefore, that increasing barring density represents the evolutionarily accessible path between a pale and a dark underwing — a way of progressively darkening the gestalt appearance of the wing through gradual quantitative shifts, until a separate switch to uniform feather pigmentation eventually occurs through one or more additional mutations (note that, for instance, variegatus shows a barring thickness similar to that of hudsonius; but in the latter, the underwing feathers are already uniformly brownish/buff). The clinal, graded nature of the geographic pattern described in this post — from white to barred to dark — is precisely what one would expect from gradual, geographically structured selection acting on this kind of developmental system, rather than on a simple on/off pigmentation switch.
A final observation: barring appears to be a common plumage feature across Scolopacidae, whereas plovers lack it altogether — and accordingly, their underwing switch seems to be more binary, jumping directly from pale to dark without the intermediate barred states that make the gradient so apparent in godwits and curlews.
What about other bird families?
The underwing may play a broader role in guiding bird migration than is currently appreciated, well beyond shorebirds. In thrushes, for instance — species that spend much of their lives concealed within woodland but undertake open-country nocturnal migration — many species display surprisingly distinctive underwing patterns that could serve as visual cues during migratory flight. The Eurasian migratory Turdus complex is instructive here: Song Thrush, Redwing, and Fieldfare each show characteristic and species-specific underwing coloration, rarely visible on the ground but fully exposed in flight.
A complementary perspective comes from ducks. The Northern Pintail, which is essentially uniform across its entire Holarctic range with no distinct subspecies or isolated populations, shows striking intraspecific variation in underwing pattern within a single population — individuals with clean white and heavily barred underwings occur side by side, apparently at random. This suggests that underwing coloration in Pintail has not been subject to any consistent directional selection, and the variation is essentially neutral. The Wigeons offer a revealing contrast: the Eurasian and American Wigeons have diverged into two distinct species, and one of their most consistent and diagnosable differences is precisely their underwing — pale in American, distinctly darker in Eurasian. Whether underwing divergence was a driver of that speciation or merely a byproduct of allopatric isolation remains impossible to assess causally — but the contrast with the Pintail situation is suggestive. Where selection on underwing pattern is absent, variation remains unstructured; where it has acted, it may have contributed to drawing a line between species.
Conclusions
This post has described a previously unreported global pattern of geographical variation in the underwing coloration of large shorebirds, consistent across multiple independent lineages and unlikely to have arisen by chance. We have proposed that shared migratory flyways — and the mixed-species flocks they generate — may represent the selective context driving this convergence, drawing a loose parallel with the acoustic convergence documented in migratory warblers by Gayk et al. (2021). We have also highlighted a number of intriguing individual cases — the Nearctic origin of islandica Knots, the position of baueri Bar-tailed Godwits at the crossroads between Pacific and Nearctic migratory systems, and even the speciation of the Wigeon pair — that seem to fit neatly within this framework, even if causality remains impossible to demonstrate. It should be emphasised that everything proposed here is highly speculative: the pattern itself still needs to be quantified rigorously, the proposed mechanism has not been tested, and alternative explanations have not been ruled out. There is much left to understand — but hopefully this is a good starting point for discussion, and perhaps an incentive for someone to continue exploring these ideas.
Acknowledgements
I would like to thank Martí Franch for discussing this post with me and helping to improve it in several ways. I am also grateful to all the photographers who kindly allowed me to reproduce their wonderful images here.
References
Brooke, M. de L. (1998). Ecological factors influencing the occurrence of 'flash marks' in wading birds. Functional Ecology, 12: 339–346.
Buehler, D.M., Baker, A.J. & Piersma, T. (2006). Reconstructing palaeoflyways of the late Pleistocene and early Holocene Red Knot Calidris canutus. Ardea, 94(3): 485–498.
Conklin, J.R., Verkuil, Y.I., Battley, P.F., Hassell, C.J., ten Horn, J., Johnson, J.A., Tomkovich, P.S., Baker, A.J., Piersma, T. & Fontaine, M.C. (2022). Global flyway evolution in red knots Calidris canutus and genetic evidence for a Nearctic refugium. Molecular Ecology, 31(7): 2124–2139.
Gayk, Z.G., Simpson, R.K. & Mennill, D.J. (2021). The evolution of wood warbler flight calls: species with similar migrations produce acoustically similar calls. Evolution, 75(3): 719–730.
Ho, W.K.W. et al. (2019). Feather arrays are patterned by interacting signalling and cell density waves. PLOS Biology, 17(2): e3000132.
Withrow, J.J. & Winker, K. (2014). Genetics of a cryptic speciation event within the golden plover complex. Wilson Journal of Ornithology, 126(3): 429–442.